Understanding the five
WH questions of Linkage
Dr. Tirth Raj Ghimire
Faculty of Science, Nepal Academy of Science and Technology, Khumaltar, Lalitpur, Nepal. Email:
ghimiretr@hotmail.com
“..it is necessary, especially at the secondary level, to
give an accurate view of science, not an oversimplification. To present the
good sides of science, and the less glorious ones. To give a fair report of
history, not a childish or idealistic view. To show that scientific truth is
not the ultimate truth, but one way of seeing things amongst others. To be a
bit less assertive in our teaching… and more open-minded to the reality of
science today and in the past, could lead us to being much more efficient”[1].
Background of this
paper
I was listening to the lecture given by my one of the best teachers, late Mr.
Bhagwan Ghimire about cell division. He was introducing meiosis cell division[2] and
elaborating how gametes of diverse characters are produced after crossing-over[3] in
meiosis. At the same time, I was curious to know what would happen if there
were the absence of crossing-over. To my curiosity, Mr. Ghimire replied that
genes would be stayed together and would be carried in the gametes without
change. He added that it would be called linkage and it would result in the
transmission of characters that is carried by gametes containing genes from
either father or mother.
I did not understand what Mr. Ghimire meant for linkage. I again asked him why
the genes do not undergo crossing-over. He replied that he would prefer to get
me read the five WH questions (what is linkage and why, where, when and how does linkage occur?) after I went to the
university. He added that after I study the history of Mendel[4], Bateson[5] and
Morgan[6], I would have had the substantial
knowledge of linkage and crossing-over. Unfortunately, he
was overwhelmed by the Narayani River after a few days, when he went to swim in
a sunny day. He never returned but his saying about the concept of absence of
crossing-over hugely stroked my scientific mind. Therefore, I consulted
the modern-day scientists and their literatures related to the Three Scientists
to understand about the five WH questions. In my view, Mr. Ghimire is one of
the rarest teachers in my secondary and higher secondary level where I got an
accurate idea of science as stated in the meeting in 2001 when Heudier
discussed the role of history and philosophy in physics education in Europe.
What is linkage and when, why, where and how does it
occur?
Linkage is the tendency of a
pair of contrasting characters or genes[7](allele[8]) to be inherited together during meiosis
cell division. This is in contrast to the principle of the Law of Independent
Assortment discovered by Mendel after his studies in pea plants (Pisum
sativum). This principle states that different genes independently separate
from one another during the development of gametes (spermatogenesis[9] or
oogenesis[10]). Mendel founded this principle because
the combinations of traits or characters in the offspring of his crosses did
not always match the combinations of traits in the parental organisms. So, the
main feature of this law is crossing-over or recombination that occurs during
meiosis. Notably, recombination occurs in two phases: firstly, breakage
of DNA[11] takes
place. Breakage secures the independent separation of genes from each other.
Secondly, the broken parts of maternal and paternal DNA (genes) are recombined
to produce new combinations of genes[12]. As a result, gametes get new
combinations of characters from the offspring. However, in some situations like
in linkage, recombination
either partly is taken place or is completely absent. So, linkage occurs during gametogenesis allowing
inheritance of linked genes.
Linkage occurs because of the nearly located
genes (linked genes) and the absence of crossing-over of chromosomes. We know
that as a result of recombination, the production of new combination of various
genes (characters) will be secured in the reproductive cells (sperms or ova) in
an individual; remarkably, the characters will be different from his/her father
and mother. In contrast, if a gamete is produced without recombination in an individual,
the gamete may carry either of his/her mother’s or father’s genes. This may
occur when genes are located very close to one another on the same chromosome
and these genes fail to undergo crossing-over and recombination. This situation
is called linkage in which the law of independent
assortment is not followed by these genes.
Who found linkage?
“Bateson, Punnett and
Edith Saunders around 1906, when making crossing experiments with sweetpeas,
realised that some characters were inherited together, in disagreement with
Mendel’s principle of independent segregation (Bateson, Saunders & Punnett
1906). At the outset they only described the phenomenon, without trying to
explain it, and they called it coupling. In 1911 they tried to explain it
through a special cell division process (reduplication hypothesis). This
hypothesis was independent of the chromosome hypothesis. The same phenomenon
was later called linkage by Morgan and his associates[13].”
In 1905, William Bateson, Edith Rebecca Saunders[14],
and Reginald Punnett[15] conducted an experiment that involved
the dihybrid crosses in a fashion similar to that carried out by Mendel. The
contrasting characters in homozygous[16] sweetpea
plants (Lathyrus odoratus) were Purple Flowers and Long Pollen grains
versus red flowers and round pollen grains. They had already noted that Purple
Flowers (P) were dominant[17] over Red flowers (p), and that Long
Pollen grains (L) were dominant over Round pollen grains (l). Notably, they
found the heterozygous plants (PpLl) that had Purple Flowers and Long
Pollen grains in F1 generation[18]. They again crossed the F1 plants with each other (between
homozygous pea plants with Purple Flowers and Long Pollen grains and homozygous
pea plants with Red Flowers and Round Pollen grains). However, they got the
ratio 14:1:1:3.5 (1528 plants with Purple Flowers and Long Pollen grains, 106
Purple Flowers and Round Pollen grains, 117 Red Flowers and Long Pollen
grains and 381 Red Flowers and Round Pollen grains in a fashion different from
those obtained by Mendelian Dihybrid Cross[19] (9:
3: 3: 1). In another part, following test-cross[20], they observed 7:1:1:7 ratio in F2 generation instead of Mendelian Test
Cross Ratio (1:1:1:1) in coupling[21]phase (See
FIGURE 1). In the same way, when two such dominant alleles or two recessive
alleles came from different parents, they tended to remain separate in
repulsion[22] phase (See FIGURE 1) with the ratio of 1:7:7:1 instead
of Mendelian Test Cross Ratio. Observing these results, they could not explain
this difference on the basis of chance alone. As they got high frequency of
parental phenotypes[23], they thought that it would be due to a
coupling or connection between the dominant alleles P and L and between the
recessive alleles p and l. They couldn’t interpret their results in terms of
the behavior of genes located on the same chromosome. This concept of why
certain alleles are coupled or linked was further explained by Thomas Hunt (TH)
Morgan.

{kind=link}
TH Morgan firstly explained the relation of linkage to the segregation of homologous
chromosomes and the occurrence of crossing-over between homologous chromosomes
during meiosis. In 1911, this explanation of linkagewas
published in a paper containing his results of crosses involving linked genes
in the fruit fly Drosophila.
Notably, he and others like Bridges[24], Muller[25], and Sturtevant[26] are
credited for most of the current theories of linkage,
crossing-over and chromosome mapping. They explained that the coupling and
repulsion jointly constitutes a larger phenomenon called linkage. Their experiments
suggested that genes were real objects, could be inherited, undergo
recombination and could be mapped to specific locations on chromosomes.
Consequently, Punnet explained linkage groups in his previous plants
(sweetpea) in 1923 and 1927. Finally, Morgan and Sturtevant detailed gene
location and gene mapping by conducting various experiments.
The story of how TH Morgan discovered linkage is interesting. In 1910, he used
various environmental and nutritional factors such as heat, cold, X-rays,
acids, bases, sugars and other chemicals and discovered a single white
fruitfly,Drosophila melanogaster. He crossed White-eyed Male Fly to its
Red-eyed sisters. Interestingly, when Morgan later inbred the heterozygous F1 Red-eyed
flies, the traits of the F2 progeny did not assort
independently. This was because Morgan observed 2,459 Red-eyed Females, 1,011
Red-eyed Males, 782 White-eyed Males, 0 White-eyed Female instead of 1:1:1:1
ratio. He again conducted another cross between the original White-eyed Male
Fly and F1 Females and
observed 129 Red-eyed Females, 132 Red-eyed Males, 88 White-eyed Females and 86
White-eyed Males. These results led him to analyze that the eye-Color was
related to the sex factor and hypothesized that the two traits were linked
together. Notably, in Drosophila,
the Gray body Color (B) is dominant over Black (b) and Long wings (V) over
Vestigial wings (v). Morgan (1910) crossed a Gray-Vestigial Male (BBvv) with a
Black-Long Female (bbVV), and obtained Gray-Long (BbVv) offspring in F1 generation. When F1 Males (BbVv) were test crossed with
Black-Vestigial Female (bbvv), only two types of F2 offspring appeared, i.e., Black-Long
(bbVv) and Gray-Vestigial (Bbvv) indicating complete linkage in Drosophila males. In the same way, when the F1 Female Fly was test crossed with a
Black-Vestigial Male, the F2 offspring
appeared in the phenotypic ratio of 8.5% Gray-Long, 41.5% Gray-Vestigial, 41.5%
Black-Long and 8.5% Black-Vestigial, i.e., the parental combinations were 83%
and recombination only 17%. Again, Black-Long was crossed with Gray-Vestigial
individuals, F1 (Gray-Long)
was test crossed, the F2 ratio
was 41.5% Black-Long, 41.5% Gray-Vestigial, 8.5% Gray-Long, 8.5%
Black-Vestigial, i.e., the parental combinations were 83% and recombination was
17% indicating incomplete linkage in these cases.
Another example of incomplete linkage comes from the work of CB Hutchinson
(1922) who took the maize of two characteristics, Colored Aleurone (C) or
Colorless Aleurone (c) and Full Endosperm (S) or Shrunken Endosperm (s). In
coupling phase, he crossed a dominant homozygous Colored Full (CCSS) with a
recessive Colorless-Shrunken (ccss) plant. The F1 plant produced Colored-Full grains
(CcSs). On test crossing with F1 (Colored-Full
grains, CcSs), the F2 offspring
were produced as parental combinations: 4032 CcSs and 4035 ccss and
recombinations: 149 Ccss and 152 ccSs. This gave the ratio of the parental
combinations and recombinations in 96.4% and 3.6% pattern despite the Mendelian
test cross ratio of 25%:25%:25%:25%. In the second phase (repulsion phase), he
crossed ccSS with CCss and obtained Colored-Full (CcSs) offspring in F1 generation. Test crossing with F1,
he obtained F2 offspring
as parental combination 97.06% (21379 CcSs and 21906 ccSs) and recombinations
2.94% (638 CcSs and 672 ccss) proving the occurrence of incomplete linkage.
What are the
historical examples of linkage experiments?
Various literatures show scientists have already discovered incompletelinkage.
For example, Bateson, Saunders and Punnet (1906) reported 7:1:1:7 in sweetpea
after crossing plant between Blue Factor[27] and Long Pollen and Red Color and
Round Pollen. Similarly, these groups in 1908 reported 15:1:1:15 ratio in the
same plants. In the same year, they reported the same ratio as 15:1:1:15 in the
sweetpea (Dark Axil and Fertility vs Light Axil and Sterility). In 1911, de
Vilmorin and Bateson crossed pea plants (Tendril and Round Seed with plants no
Tendril or Acacia type and Wrinkled Seed) and found a
ratio of 63:1:1:63. Gregory in 1911 reported 7:1:1:7 ratio in the plants
(Magenta Color and Short Style with the Red Color and Long Style). Bateson and
Punnet in 1911 reported the ratio of 127:1:1:127 after crossing the sweetpea
(Blue Factor and Erect Standard with the Red Color and Hooded Standard). There
are several examples of these types of gametic coupling in literatures. In the
same way, Bateson, and Punnet in 1911 found 1:3:3:1 ratio while crossing
between sweetpea (Normal Flower and Fertility with the ‘Cretin’ type and
Sterility). Both scientists also reported 1:7:7:1 ratio after crossing between
the plants with Blue Factor and Long Pollen and Red Color and Round
Pollen.
Several scientists reported the complete linkage in plants and animals. For example,
Emerson in 1911 reported this phenomenon in maize (Dark Purple Husks and
Purplish Pericarp vs White Husks and Colorless Pericarp or Red Cob and Red
Pericarp vs White Cob and Colorless Pericarp). Bateson and Punnet in 1911
reported this phenomenon in sweetpea (Dark Axil and Fertility vs Light Axil and
Sterility). Bateson, Saunders and Punnet in 1908 reported complete or very high
intensity in sweet pea (Erect Standard and Blue Factor vs Hooded Standard and
Red Color). Baur in 1912 reported complete linkage in Aquilegia(Variegated
Green leaf and Homogenously Green leaf vs Absence of ‘Variegate’ Factor and
Absence of Homogeneously Green Factor, i.e., ‘Chlorina’ Color). Correns in 1912
also reported this phenomenon in Silene
Armeria (Rich Pigment and
Saturator and Less Pigment and Absence of Saturator). Raynor and Doncaster in
1906 and Doncaster in 1908 reported the similar phenomenon inAbraxas (Currant Moth) (Femaleness and
‘Grossulariata’ factor vs Maleness and LactiColor Character).
How do you calculate linkage map distance?
One linkage map unit (LMU or linkage map distance) is defined as 1%
recombination (% recombination between genes) or recombinant gametes or
phenotypes. It means a map unit is equivalent to the physical distance along a
chromosome that will experience 1 crossover event in every 50 meiotic divisions.
In this context, 1 crossover in 50 meiotic divisions is equal to 2 recombinant
gametes in every 200. This is equal to 1% recombination. Therefore, two genes
that recombine with a frequency of 1% are said to be 1 map unit apart. One map
unit is also called as one centimorgan (cM) (1cM = 1 megabase (Mb) = one
million nucleotides).
To calculate the distance between any two genes, the number of recombinant
gametes is divided by the total number of gametes. These are just estimates and
the difference between the two estimates reflects random deviation. Notably,
these estimates are not incorrect, as accuracy depends on the replicates and
huge data. However, LMU do not correspond to any fixed length of chromosome.
Firstly, frequency of crossover and recombination can be affected by location.
This is because crossing-over is inhibited near the centromere and immediate to
the point of another cross-over. Secondly, double cross-overs are usually
missed because the number of recombination events is always
underestimated.
Let’s consider an example how to calculate the distance between any two linked
genes, for example the genes that determine Color (C) and Endosperm (S) by the
help of Hutchinson’s experiment in maize. In the coupling experiment,
Total offspring =
4032+4035+149+152= 8368.
Total recombinant =
149+152= 301.
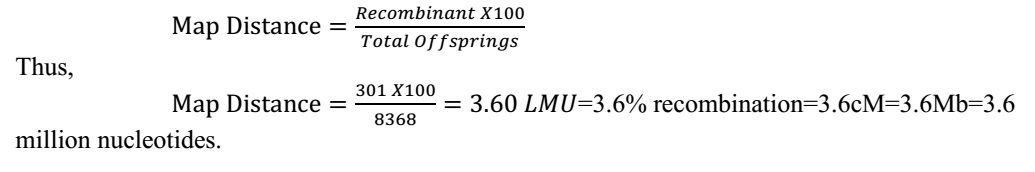
Interestingly, scientists believe that non-crossovers (linkage) vary
from 50% to 100%. 100% non-crossovers (complete linkage, Recombination Frequency=0)
is a state where no crossing-over takes place as observed in maleDrosophila.
In the same way, the cross-over ranges from 0 to 50% (Recombination
Frequency=0.5) but never exceeds 50%. These values are true when genes are
on different chromosomes or genes are separated very far apart on the same
chromosome. A recombination frequency of 0.5 indicates that 50% of offspring
are recombinants and the other 50% are parental types. So, every type of allele
combination is represented with equal frequency.
What is the
significance of linkage?
Firstly, linkage holds the parental characters
together. Secondly, it restricts the appearance of new recombination or new
characters. Thirdly, marker genes such as Number of flowers, Weight of seeds,
Lint Length that are linked to some quantitative characters are economically
important for plant breeders. For example, the Corolla Color is marker for lint
index in cotton. The grain Color is marker for the yield of rice.
Acknowledgement: I acknowledge late Mr. Bhagwan Ghimire for his well discussion about the
mechanisms of crossing-over and meiotic events in the eukaryotic cells during
my schooling (grade 9). I am grateful to various scientists around the globe
for the discussion of the linkage phenomenon and various online
materials referred in this text (Accessed on 1st October 2013).
Footnotes
[1] Heudier, J.-L.:
2001, ‘The role of history and philosophy in Physics Education’, in H. Wilson
and B. Warmbein (eds.), Physics on Stage. Full Proceedings 2000.
European Space Division, Noordwijk, 22-23.
[2] A type of cell
division in which a diploid (two set of chromosome) cell divides into four
haploid (one set of chromosome) cell.
[3] An exchange of
genetic material between homologous chromosome.
[4] Gregor Johann
Mendel (July 1822– January 1884), a German-speaking Austrain scientist,
was the Founder of the modern Genetics. He discovered the laws of inheritance.
[5] William Bateson
(August 8, 1861–February 8, 1926), a British Geneticist and a Fellow of St.
John’s College, Cambridge was the first person to use the term genetics to
describe the study of heredity and biological inheritance.
[6] Thomas Hunt Morgan
(September 25, 1866 – December 4, 1945) was an American Evolutionary Biologist,
Geneticist and Embryologist. He was awarded Nobel Prize in Physiology or
Medicine in 1933 because he explained the role of chromosomes in heredity.
[7] A unit of genetic
information that occupies a specific position on a chromosome and comes in
multiple versions called alleles.
[8] One alternative of
a pair or group of genes that could occupy a specific position on a chromosome.
[9] The development of
male gamete (sperm)
[10] The development of
female gamete (ovum).
[11] Deoxyribonucleic
acid; the molecule in which genetic information is encoded or written.
[12] Gene is a unit of
genetic information that is present in a specific position on a DNA in higher
organisms like from protista to mammalia kingdom or monera to Angiospermiae
kingdom or DNA or RNA in lower organisms like viruses.
[13] Martins, L. A.-C.
P.: 1997, A Teoria Cromossômica da Herança, Proposta, Fundamentação,
Crítica e Aceitação, Unicamp, Campinas, Ph.D. thesis.
[14] Edith
Rebecca Saunders (1865–1945), a British Geneticist and Plant Anatomist and a
Fellow of the Royal Horticultural Society, rediscovered Mendel’s laws of
heredity.
[15] Professor Reginald
Crundall Punnett (June 1875–January 1967), a British Geneticist and
Professor of Biology at Cambridge, created the Punneett Square and
wrote ‘Mendelism’, the first popular textbook on genetics.
[16] With a genotype
with two of the same alleles for a trait or character.
[17] An allele that
masks a recessive trait and may produce the same the same phenotypic effect
whether it is inherited as heterozygous or homozygous.
[18] The first
offspring generation or the first filial generation of seeds or plants or
animal offspring resulting from the cross mating of distinctly different
parental types.
[19] Dihybrid cross
involves the inheritance of two types of traits of an organisms. This theory
was invented by Mendel to determine if different traits of pea plants, for
example, the flower Color and seed shape were inherited independently.
[20] The test-cross shows how traits are inherited when the offspring of the
testcross shows a certain phenotype. If the offspring shows the recessive
trait, it indicates the heterozygous parent (F1). If the offspring
are all dominant, then it indicates the homozygous F1.
Test cross is also used to calculate the ratio between non-crossover and
crossover.
[21] When the two
dominant genes are present on the chromosome and their recessive alleles on the
other chromosome of the homologous pair, the linkage relationship
is called coupling.
[22] When one dominant
and one recessive allele are present on each chromosome of the homologous pair,
the linkage relationship is called repulsion.
[23] The physical or
observable characteristics of an organism.
[24]Calvin Blackman
Bridges (January 1889 – December 1938), an American geneticist, was one of
the scientists of the Fly Room of TH Morgan at Columbia University and wrote
the Ph.D. thesis on “Non-disjunction as proof of the chromosome theory of
heredity.”
[25] Hermann Joseph
Muller (HJ Muller) (December 21, 1890 – April 5, 1967) was an American
Geneticist, educator and Noble Prize laureate who got this award in 1946 for
the production of mutations by means of X-rays.
[26] Alfred Henry
Sturtevant (November 1891 – April 1970), an American Geneticist
constructed the first genetic map of a chromosome in 1913 and worked on Drosophila
melanogaster with TH Morgan.
[27] Blue factor means
the Color of flower is purple.
Bibliography
Bateson, W., et
al. Experimental studies in the physiology of heredity. Reports to
the Evolution Committee of the Royal Society 2, 1–55, 80–99
(1905)
Benson, K. R. T. H.
Morgan's resistance to the chromosome theory. Nature Reviews Genetics 2,
469–474 (2001) doi:10.1038/35076532
Bridges, C. B. A
revised map of the salivary gland X-chromosome. Journal of Heredity 29,
11–13 (1938)
Bridges, C. B.
Salivary chromosome maps with a key to the banding of the chromosomes of Drosophila melanogaster. Journal
of Heredity 26, 60–64 (1935)
Mendel, G. Experiments
in plant hybridization. Trans. by William Bateson. (1866)
MLA style:
"Thomas H. Morgan - Biographical". Nobelprize.org. Nobel
Media AB 2013. Web. 4 Oct 2013.http://www.nobelprize.org/nobel_prizes/medicine/laureates/1933/morgan-bio.html
Morgan, T. H. (1910).
Random segregation versus coupling in Mendelian inheritance. Science 34,
384 (1911)
Morgan, T. H. (1910).
Sex-limited inheritance in Drosophila. Science 132,
120–122 (1910)
Pierce, B. Genetics:
A Conceptual Approach, 2nd ed. (New York, W. H. Freeman, 2006)
Punnett, R. C. Linkage groups and chromosome number in Lathyrus. Proceedings
of the Royal Society of London: Series B, Containing Papers of a Biological Character 102,
236–238 (1927)
Punnett, R. C. Linkage in the sweetpea (Lathyrus odoratus). Journal
of Genetics 13, 101–123 (1923)
Sadava, D., et
al. Life: The Science of Biology, 8th ed. (New York, W. H.
Freeman/Sinaeur Associates, 2008)
Sturtevant, A. H. The
linear arrangement of six sex-linked factors in Drosophila, as
shown by their mode of association. Journal of Experimental Zoology, 14,
43–59 (1913)
Please refer this
article by properly writing author's name, title, and date of publish of this
manuscript (Ghimire TR. Understanding the five WH questions of Linkage.
Online Blog: www.hamrobato.blogspot.com, published in 20 July, 2016). It
is not allowed to use this article without permission from author (Please ask
Author at ghimiretr@hotmail.com before using it! Thanks!